All published articles of this journal are available on ScienceDirect.

Changes in Midbrain Pain Receptor Expression, Gait and Behavioral Sensitivity in a Rat Model of Radiculopathy
Authors Info & Affiliations
Abstract
Intervertebral disc herniation may contribute to inflammatory processes that associate with radicular pain and motor deficits. Molecular changes at the affected dorsal root ganglion (DRG), spinal cord, and even midbrain, have been documented in rat models of radiculopathy or nerve injury. The objective of this study was to evaluate gait and the expression of key pain receptors in the midbrain in a rodent model of radiculopathy. Radiculopathy was induced by harvesting tail nucleus pulposus (NP) and placing upon the right L5 DRG in rats (NP-treated, n=12). Tail NP was discarded in sham-operated animals (n=12). Mechanical allodynia, weight-bearing, and gait were evaluated in all animals over time. At 1 and 4 weeks after surgery, astrocyte and microglial activation was tested in DRG sections. Midbrain sections were similarly evaluated for immunoreactivity to serotonin (5HT2B), mu-opioid (µ-OR), and metabotropic glutamate (mGluR4 and 5) receptor antibodies. NP-treated animals placed less weight on the affected limb 1 week after surgery and experienced mechanical hypersensitivity over the duration of the study. Astroctye activation was observed at DRGs only at 4 weeks after surgery. Findings for pain receptors in the midbrain of NP-treated rats included an increased expression of 5HT2B at 1, but not 4 weeks; increased expression of µ-OR and mGluR5 at 1 and 4 weeks (periaqueductal gray region only); and no changes in expression of mGluR4 at any point in this study. These observations provide support for the hypothesis that the midbrain responds to DRG injury with a transient change in receptors regulating pain responses.
INTRODUCTION
Herniation of the intervertebral disc (IVD) is a common cause of low back pain and may be associated with nerve root dysfunction, known as radiculopathy. Features of radiculopathy can range from radiating neuropathic pain to paresthesias and sensory deficits to motor deficits, and the societal cost of this pathology involves over 660,000 hospital visits and over 325,000 surgeries for discectomy procedures annually in the USA [1]. The consequent mechanical compression and biochemical irritation of nearby neural structures seems to be mediated by heightened inflammatory and immune cytokine expression alongside inflammatory cell infiltration [2-7]. Indeed, upon systemic exposure, the nucleus pulposus (NP) component of IVD tissue initiates a specific immune response, likely a consequence of its immune privileged avascular location bounded by the anulus fibrosus (AF) [8, 9].
There is clear interest in identifying the cell populations affected by disc herniation-induced radiculopathy, and the role of neurotransmitters and their receptors that mediate the symptomatic and functional deficits of radiculopathy. Animal models based on compression injury of the dorsal root ganglion (DRG) demonstrate resultant allodynia and functional deficits in association with an elevated microglial activation in the spinal cord [10-14]. In addition, molecular and mRNA changes occur locally at the DRG or in the spinal cord implicating key inflammatory mediators (e.g., TNF-α, IL-1, nitric oxide) and neuropeptides (e.g., substance P) following DRG compression injury or exposure to autologus NP [9, 15-23]. These observations suggest a hypothesis that neuroplastic remodeling of the spinal cord can contribute to or modulate the pain and symptoms of disc herniation-associated radiculopathy.
The goal of the current study was to expand the scope of the disc herniation associated radiculopathy model to understand not only the induced pain phenotype, but also the mechanisms underlying spontaneous functional recovery. In prior studies of a model of non-compressive disc herniation with autologous NP material, animals exhibited behavioral changes including heightened behavioral mechanical sensitivity, stance symmetry, and gait parameters including symmetry and force analysis. Divergent findings of gait symmetry andmechanical sensitivity suggested supraspinal modulation of the nociceptive phenotype and promoted investigation into mechanisms underlying that descending control. An important objective of the current study was to evaluate a role for modified neurotransmitter receptor expression in the central nervous system at the midbrain to test if neuroplastic remodeling can occur central subsequent to DRG injury or sensitization. Thus, transient changes were studied here in a rodent model of non-compressive disc herniation. Immunohistochemical markers of neuroinflammation at the DRG and the presence of a monocyte chemotactic protein (MCP-1) were quantified in the cerebrospinal fluid. The expression of serotonin, metabotropic glutamate, and mu-opioid receptors was characterized in the periaqueductal gray and red nucleus regions of the midbrain by immunohistochemistry.
MATERIALS AND METHODOLOGY
Animal Model
Male Sprague-Dawley rats (n=30, 9 months old) were divided into sham (n=12), NP-treated (n=12), and pre-operative control (n=6) groups. The sham and NP-treated groups underwent a previously described operation [15] as approved by Duke University Institutional Animal Care and Use Committee (IACUC) while the pre-operative control animals served as non-surgical control tissue.
In brief, under pentobarbital anesthesia, rats underwent surgical exposure of proximal tail IVD in which a single disc was punctured, the NP was harvested via curette, and the tail was closed. In these same animals, a dorsal exposure of the right L5 DRG was accomplished by a partial unilateral laminotomy and medial facetectomy, with autologous NP either discarded (sham) or placed onto the exposed DRG (NP-treated). Two-layer closure was performed and animals were returned to solitary cage housing for 5–7 days, followed by group housing until sacrifice.
Animals in sham and NP-treated groups were studied longitudinally for gait, stance symmetry, and behavioral sensitivity at pre-operative and post-operative time points. Animals were then sacrificed at 7 or 28 days post surgery (n=6 per animal group for each time point) after which the cerebrospinal fluid, midbrain and DRG were harvested. The pre-operative control animals were procured and midbrain harvested only to serve as a control group.
Geometric and Temporal Gait Descriptors
To assess rodent gait, rat ambulation was recorded while freely exploring a custom-built gait arena (5’6” x 1’6”) pre-operatively and at 1 and 4 weeks after surgery, as previously described [15]. When a rat passed through the middle of the arena, a high-speed video camera was manually triggered to capture the rat’s movement (Phantom V4.2, 200 frames per second; Vision Research, Wayne, NJ). Trials that contained a minimum of two complete gait cycles and a consistent velocity were saved for processing (5 trials per animal). Using custom code (DLTdataviewer), the video frame of foot-strike and toe-off events were determined viaby-hand digitization [24, 25]. Using these data, percentage stance time imbalance and gait symmetry were calculated for each trial. Percentage stance time (also known as limb duty factor) is the percentage of time a limb is in stance for a given stride, or mathematically as stance time divided by stride time [26]. Gait symmetry is the offset between left and right foot-strikes in a limb pair for a given stride, or mathematically as the time between left and right foot-strikes divided by time between two left foot-strikes [26]. Rats commonly ambulate with balanced, symmetric gaits, represented mathematically by a difference between the left and right percentage stance times of 0 and a gait symmetry of approximately 0.5; a shift in these variables would indicate an unbalanced or asymmetric gait, respectively. Data for each group (sham, NP-treated) were compared to the mathematical definitions for balanced, symmetric gait using a repeated measures t-test on time with a Bonferroni correction; differences amongst treatment groups and between pre-operative and post-operative timepoints were analyzed using a two-way ANOVA with a post-hoc Tukey’s test.
Weight Distribution
Hind limb weight distribution was determined preoperatively and at 1 and 4 weeks after surgery. An incapacitance meter (IITC, Inc.) was used to measure weight distribution on the left and right limb, and data was acquired during 5 second intervals (5 trials per rat). Weight distribution for each group was compared to the mathematical definitions for balanced weight distribution using a repeated measures t-test on time with a post-hoc Bonferroni correction (unbalanced distribution ≠ 50% weight on right limb); differences amongst groups (sham, NP-treated) and between pre-operative and post-operative timepoints were analyzed using a two-way ANOVA.
Mechanical Sensitivity
Mechanical paw withdrawal thresholds were determined preoperatively and at 1 and 4 weeks after surgery using an up-down protocol described by Chaplan and coworkers [27]. Briefly, rats were placed in a wire-bottom cage and von Frey filaments (Stoelting) were applied to the plantar surface of rat’s hind paws. If paw withdrawal was observed, the next smallest filament was applied; if paw withdrawal was not observed, the next largest filament was applied. Using this up-down protocol, the 50% paw withdrawal threshold can be approximated; this threshold represents the mechanical force where paw withdrawal and stimulus tolerance are equally likely. A two-factor ANOVA followed by a post-hoc Tukey’s test was used to compare differences amongst treatment groups (sham, NP treated) and between pre-operative and post-operative timepoints for the affected limb.
DRG Immunostaining
At the time of dissection, lumbar DRGs from sham and NP-treated groups were dissected en bloc and placed in optical cutting temperature (OCT) embedding media (Sakura Finetek, Torrence CA) and frozen in liquid nitrogen until sectioning. Cryosections of DRG (7μm) were immunostained for glial fibrilary acidic protein (GFAP) to identify astrocytes. DRG sections were fixed in cold acetone (10 min, -20°C), permeabilized with 0.02% Triton-100, and incubated with blocking serum (30 min, 3.75% BSA (Gibco) and 5% goat serum (Zymed, Invitrogen, Carlsbad, CA)) to minimize non-specific antibody binding. Sections were incubated with primary monoclonal mouse GFAP antibody (GFAP, SC-51908, Santa Cruz Biotechnology, Santa Cruz, CA) diluted 50x in blocking serum (2 hours at room temperature), washed with PBS and incubated with Alexa Fluor 488-conjugated secondary antibodies (30 min at 200x dilution in blocking serum, Molecular Probes, Invitrogen).
Separate DRG sections were immunostained to detect the presence of ionized calcium binding adaptor molecule 1 (Iba1) to determine presence of macrophages. DRG cryosections were fixed in cold methanol and acetone mixture (1:1) for 5 min ( -20°C), permeabilized with 0.5% Triton-100 (15 min, room temperature) and incubated with blocking serum (30 min, 1% BSA, 0.25% Triton-100, & 5% goat serum). Sections were incubated with rabbit polyclonal Iba1 antibody (Wako, 019-19741) with a 250x dilution in blocking serum overnight. DRG sections were washed with PBS and incubated with Alexa Fluor 488-conjugated secondary antibodies (30 min at 200x dilution in blocking serum, Molecular Probes).
All stained DRG sections were counterstained with 0.5 mg/ml propidium iodide (Sigma-Aldrich) at room temperature for 15 minutes to stain the cell nuclei and mounted with GVA mounting medium (Zymed, Invitrogen).
Stained tissue sections were imaged using a confocal laser scanning microscope (Zeiss LSM 510, 20x objective; Zeiss, Jena, Germany) with 20x magnification. Multiple digital images (526x526) were acquired for each DRG sample and evaluated by two blinded graders coming to consensus for fluorescence staining uniformity and intensity with the following ordinal scale: 0 (no stain), 1 (moderate, non-uniform stain), 2 (moderate, uniform stain), 3 (intense stain) for each image.
Grades of immunohistochemical staining for GFAP and Iba1 were tested for evidence of differences between 1 and 4 weeks time period for sham and NP-treated animals using Kruskal-Wallis one-way analysis of variance with a 0.05 level of significance. When significance was observed, post-hoc Wilcoxon Rank Sum Tests were conducted to detect differences in time for each group.
Midbrain Immunostaining
At the time of sacrifice, midbrains from all animals were dissected en bloc and placed in optical cutting temperature (OCT) embedding media (Sakura Finetek) and frozen in liquid nitrogen until sectioning. Cryosections of midbrain (25μ m) were fixed in 4% formaldehyde (10 min, room temperature), permeabilized with 0.2% Triton (10 min), and incubated with blocking solution (30 min in 3.75% BSA and 5% goat serum, Invitrogen). Sections were incubated with monoclonal mouse serotonin receptor 2b (5HT2b, 556334, BD Biosciences Pharmingen, San Jose, CA ) diluted 100x in blocking solution for 60 minutes. Samples were then washed with PBS and incubated with an AlexaFluor 488-conjugated secondary antibody (Molecular Probes) diluted 100x in blocking buffer for 30 minutes. Cell nuclei were counterstained using 0.5mg/ml propidium iodide (20 min, Sigma-Aldrich) and slides were mounted with GVA mounting solution (Invitrogen).
A separate protocol was used to immunolabel midbrain sections with antibodies against metabotropic glutamate and μ-opoid receptors as described here. Cryosections of midbrain (25 μm) were fixed in 10% formaldehyde (30 min, room temperature), quenched in peroxo-block (2 min, 1 part 30% H2O2: 9 parts methanol), and incubated in ready-to-use 2.5% normal horse blocking serum (20 min, ImmPRESS Universal Reagent kit: MP7500, Vector labs, Burlingame, CA). Samples were then incubated for 60 minutes with the following primary antibodies in blocking serum: rabbit polyclonal metabotropic glutamate receptor 5 (diluted 200x, mGluR5; ab53090, Abcam Inc, Cambridge, MA), rabbit polyclonal metabotropic glutamate receptor 4 (diluted 400x, mGluR4; 06-765, Upstate, Lake Placid, NY), or rabbit polyclonal μ-opioid receptor (diluted 100x, μ-OR, 44308G, Biosource, Camarillo CA). Samples were washed in PBS and incubated with ImmPRESS Peroxidase Universal anti mouse/rabbit Ig reagent (30 min, ImmPRESS Universal Reagent kit), washed again and incubated with diaminobenzideine-DAB solution (30 min, SK-4100, Vector labs). Cell nuclei were counterstained using Methyl Green (H-3402, Vector labs) for 5 minutes (60°C) and mounted with GVA mounting solution (Invitrogen).
Immunostained sections of the rat midbrain were imaged with a confocal laser scanning microscope (5HT2b receptor only; Zeiss LSM 510, 20x objective; Zeiss, Jena, Germany) or a light microscope (Nikon Eclipse E600 POL, 4x objective; Nikon Instruments Inc, USA). Digital images were acquired of midbrain regions and analyzed as follows. For the 5HT2b receptor, images were thresholded on red and green channels to highlight labeled cell nuclei and associated staining. The numbers of cells and positively labeled cells were counted to determine the percent of positively stained cells.
To analyze intensity of positive staining for all other receptors, the periaqueductal gray (PAG) or red nucleus (RN) regions of the midbrain were first identified. Using Matlab, 526x526 pixel sized regions within the PAG or RN were obtained and converted to grayscale images. Images were then thresholded against a corresponding negative control to highlight only positively stained cells. After thresholding, images were converted to black and white images in order to calculate percent of positively stained pixels in Matlab.
Midbrain data (% positively labeled cells or % positively labeled pixels) for each pain receptor was analyzed for differences amongst pre-operative control, shamand NP-treated groups at day 0, week 1 and week 4 time points by two-way analysis of variance (2-way ANOVA) at a 0.05 level of significance. Tukey’s post-hoc analysis was performed when significance was observed.
Cerebrospinal Fluid (CSF) Biomarker Assessment
Prior studiesdemonstrated elevated expression of monocyte-chemoattractant protein (MCP-1) in multiple sclerosis [28], acute spinal cord injury [29], and traumatic brain injury [30]. To investigate changes in CSF concentrations of MCP-1, CSF was collected at the time of sacrifice through a 23 gauge needle puncture of the 4th ventricle. Due to technical limitations, CSF samples were collected from 3 NP-treated rats at 1 week, 5 NP-treated rats at 4 weeks, 6 sham rats at 1 week, and 4 sham rats at 4 weeks. Levels of CSF MCP-1 were quantified using a commercially available ELISA (ERMCP-1C, Thermo Fisher Scientific, Waltham MA, USA) following the manufacturer’s instructions. Briefly, MCP-1 reference and rat CSF samples, diluted 1:25, were incubated in microwells coated with MCP-1 binding protein. Data were analyzed via an unbalanced two-factor, full-factorial ANOVA.
RESULTS
Mechanical Sensitivity
Mechanical allodynia was observed in the affected limb of animals exposed to autologous NP (Table 1). Lower values for 50% paw withdrawal thresholds among NP-treated animals were observed when compared to pre-operative testing, sham, and their own contralateral hindlimbs. This lower mechanical withdrawal threshold occurred early at 1 week and persisted through to the 4 week evaluation (p<0.05, Table 1). There was no evidence of a change in mechanical withdrawal thresholds in the affected limb of sham animals (p>0.05), nor was there evidence of a change in mechanical withdrawal thresholds in the contralateral limb for NP-treated or sham animals (data not shown).
Data for Gait Parameters. All Data Shown are Mean ± Standard Error
| Pre-Operative | Sham (1 Week) | Sham (4 Weeks) | NP-Treated (1 Week) | NP-Treated (4 Weeks) | |
|---|---|---|---|---|---|
| Mechanical withdrawal threshold (grams-force) | 14.6 ±1.2 | 7.4±1.1 | 11.8±2.1 | 3.4±0.7*, ⊥ | 4.3±1.0* |
| Weight-bearing (% weight on affected limb) | 49.5±1.0 | 47.3±3.0 | 50.0±0.7 | 41.9±3.1*, ∀ | 49.2±0.9 |
| Percentage stance time imbalance (% stance time, left-right limb) | 0.2±0.4 | -0.6±0.9 | 0.0±0.6 | 4.1±1.4*, ∀, # | 1.5±1.7 |
| Gait symmetry | 49.3±0.4 | 50.9±0.8 | 50.7±0.5 | 54.5±1.5*, ∀ | 52.0±2.3 |
∀ Imbalanced/asymmetric (p<0.05).
* Different from pre-operative data (p<0.05).
# Different from 1 week sham (p<0.05).
⊥ Different from 4 week sham (p<0.05).
Weight Distribution
NP-treated animals had a weight-distribution imbalance at 1 week (p <0.05, Table 1), which was significantly different from the weight distribution observed pre-surgery (p<0.02, Table 1). By week 4, balanced weight-distribution returned in NP-treated animals. Weight-distribution in sham animals remained balanced and did not vary from pre-surgery values.
Temporal Gait Descriptors
NP-treated animals used imbalanced, asymmetric gaits at 1 week (Table 1). Gait asymmetry and imbalance of limb duty factor was observed on the affected limb. Percentage stance times were significantly shorter on the affected limb relative to the contralateral limb (percentage stance time imbalance > 0%, p<0.05), and gait symmetry indicated that left and right foot-strikes were not equally spaced in time (gait symmetry ≠ 50%, p< 0.05). This temporal gait profile was significantly different from both pre-surgery values and sham controls at 1 week. By 4 weeks, NP-treated animals returned to balanced, symmetric gaits. Gait asymmetry and imbalance were not observed for sham animals.
DRG Immunostaining
GFAP expression in the DRG was found to be higherin the DRGs of NP-treatedanimals at 4 weeks post-operatively as compared to the 1-week timepoint (p<0.0004) (Fig. 1), with no evidence of differences in GFAP staining for sham animals at either timepoint. Overall, the intensity of staining was found to be very low with mean values that correspond to moderate and non-uniform staining in the sham animals.
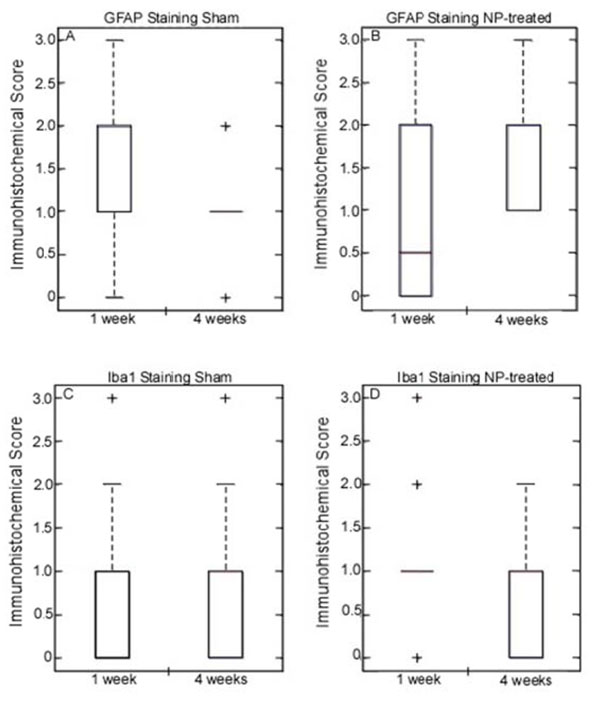
DRG Immunostaining: Glial fibrillary acidic protein (GFAP) in DRG (top; graphs A-B): Increased expression was observed in NP-treated (*p<0.0004) animals at 4 weeks after surgery. Expression levels of GFAP in sham animals remained constant at all time points. Ionized calcium binding adaptor molecular 1 (Iba1) in DRG (bottom; graphs C-D): Expression level of Iba1 did not vary in sham or NP-treated animals at any time point.
Very low levels of staining were observed in all DRGs for Iba1 expression, with no evidence of differences between 1 and 4 week timepoints in either sham or NP-treated animals (Fig. 1).
Midbrain Immunostaining
Expression of serotonin 2b receptor (5HT2b) was increased at 1 week in NP-treated animals (p<0.0015 vs unoperated controls) and returned to pre-operative control levels by 4 weeks (Fig. 2). Expression level of 5HT2b for sham animals remained the same level as pre-operative controls. Metabotropic glutamate receptor 5 expression (mGluR5) was increased in the PAG region of both sham (p<0.0001 vs pre-operative controls) and NP-treated animals (p<0.0001 vs pre-operative controls) at one week (Fig. 3). The increase in expression level of mGluR5 in the PAG at one week for NP-treated animals was significantly higher than sham animals (p<0.0001). By four weeks, mGluR5 expression returned to the same expression level as pre-operative controls in sham animals, while expression levels in NP-treated animals remained elevated (p<0.0001 vs pre-operative controls and sham animals at 4 weeks). mGluR5 expression levels in the RN of sham and NP-treated animals was not significantly different from each other and remained the same level as pre-operative controls (Fig. 3). mGluR4 expression level of sham and NP-treated animals also remained the same as pre-operative controls at all time points (Fig. 4). Increased expression of μ-opioid receptor (μ-OR) was observed after 1 and 4 weeks in sham animals (p<0.0001 vs pre-operative controls) and NP-treated animals (p<0.0001 vs pre-operative controls) as compared to pre-opreative control basal levels (Fig. 5). In NP-treated animals, expression levels of μ-opioid receptor increased even above sham animals levels at the 4 week timepoint (p<0.0001), but in PAG regions only. In the RN region, μ-opioid receptor expression levels did not change for sham or NP-treated animals (Fig. 5).
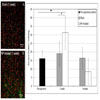
Serotonin 2b (5HT2b) Receptor Immunostaining: (left) Representative images of 5HT2b immunohistochemical images for sham (A) and NP-treated (B) 1 week after surgery (red=nuclei; green=positive staining; scale bar =100 µm). Increased expression was observed in NP-treated (** p<0.015) animals at 1 week after surgery, but returned to pre-operative levels by 4 weeks post surgery. Expression level at 1 week for NP-treated animals was significantly different from sham animals (*p <0.015). Data are presented as mean ± standard deviation.
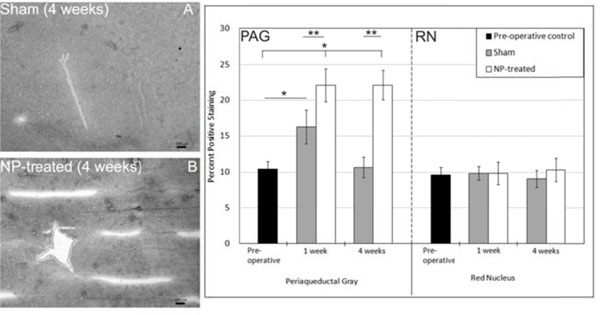
Metabotropic Glutamate Receptor 5 (mGluR5) Immunostaining: PAG: (left) Representative images of mGluR5 immunohistochemical images for sham (A) and NP-treated (B) 4 weeks after surgery (dark spots represent positive staining; scale bar =100 µm). Increased expression was observed in both sham (*p<0.0001) and NP-treated (*p<0.0001) animals at 1 week after surgery compared to unoperated controls. At one week post surgery, mGluR5 expression levels was significantly different in NP-treated compared to sham (**p<0.0001). Expression levels of mGluR5 in sham animals returned to expression levels of pre-operative controls at 4 weeks while NP-treated (**p<0.0001) animals continued to have increased expression of mGluR5. RN: Expression level of mGluR5 in the red nucleus did not vary in sham or NP-treated animals. Data are presented as mean ± standard deviation.
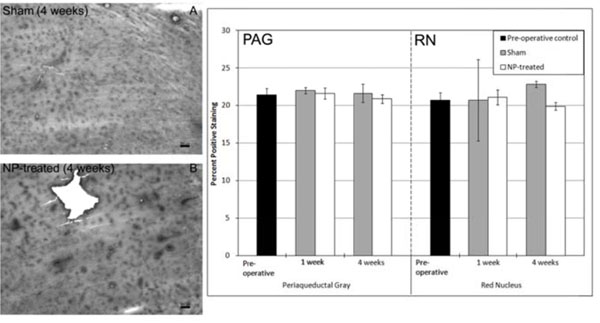
Metabotropic Glutamate Receptor 4 (mGluR4) Immunostaining: PAG: (left) Representative images of mGluR4 immunohistochemical images for Sham (A) and NP-treated (B) 4 weeks after surgery (dark spots represent positive staining; scale bar = 100 µm). Expression level of mGluR4 in the PAG did not vary in sham or NP-treated animals. RN: Expression level of mGluR4 in the RN did not vary in sham or NP-treated animals. Data are presented as mean ± standard deviation.
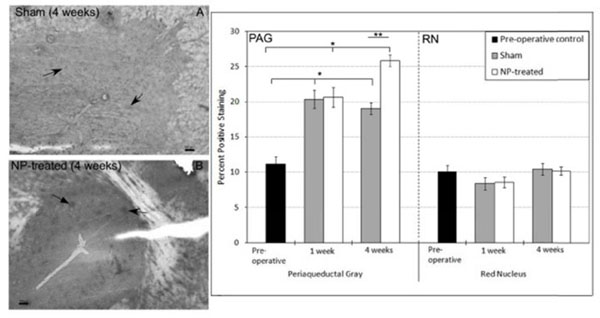
Mu-Opioid (µ-OR) Receptor Immunostaining: PAG: (left) Representative images of µ-OR immunohistochemical images for sham (A) and NP-treated (B) 4 weeks after surgery (dark spots represent positive staining; arrows indicate representative positive staining; scale bar = 100 µm). Increased expression was observed in both sham (*p<0.0001) and NP-treated (*p<0.0001) animals at 1 week after surgery. Expression levels of µ-OR in sham animals remained constant while expression levels of NP-treated animals continued to increase at 4 weeks (**p<0.0001 vs 4 week sham animals). RN: Expression level of µ-OR in the RN did not vary in sham or NP-treated animals. Data are presented as mean ± standard deviation.
Cerebrospinal Fluid (CSF) Biomarker
MCP-1 was detectable in CSF of all rats in sham and NP-treated groups at ng/ml levels (Fig. 6). When comparing MCP-1 levels between sham and NP-treated groups, MCP-1 levels tended to be higher in NP treated rats as compared to sham-operated controls at the 4 week timepoint. The magnitude of difference in MCP-1 values was large between these two groups at 4 weeks post-operatively, nearly 2-fold, although a statistically significant difference was not detected, likely due to large inter-animal variances and unequal sample numbers across groups (p=0.355, Fig. 6).
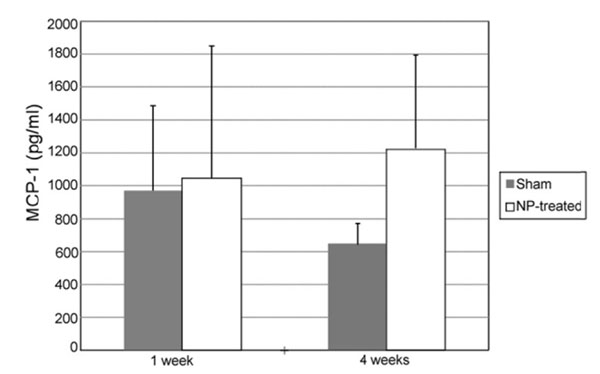
MCP-1 concentration in the CSF: At 1 Week, MCP-1 concentration was comparable between NP-treated (n = 3) and sham animals (n = 6). By week 4, NP-treated animals (n = 5) tended to have higher levels of MCP-1 relative to sham (n = 4); however, this difference was non-significant (p = 0.355). Data are presented as mean ± standard error.
DISCUSSION
Exposure of the dorsal root ganglion (DRG) to autologous nucleus pulposus (NP) has been well-studied as an animal model of radiculopathy through stab incision of the anulus fibrosus with consequent disc herniation [4], or through open placement of NP tissue upon the nerve root as performed here [13, 14]. Among these, a sustained sensitization of the DRG occurs as evidenced by elevated allodynia for the affected limb that is sustained for periods longer than 4 weeks [14]. This work similarly demonstrates mechanical allodynia in this model of radiculopathy, at both 1 and 4 weeks after NP exposure, a persistence that is incongruous with short-term changes of altered stance symmetry and impaired gait at only 1 week after surgery with subsequent normalization thereafter. Findings for elevated astrocyte reactivity, but not microglial reactivity from histological examination of the affected DRG are interesting in light of prior reports for elevated astrocytic and/or microglial reactivity in the dorsal horn of the spinal cord after DRG injury due to compression or chemical exposure [9-11], and in the DRG region itself [14]. Numerous studies suggest that spinal astrocytic and microglial activation following nerve injury depend on the mechanism of injury, and are not predictive of sustained pain-related behaviors. It is likely that supraspinal descending modulation of the pain experience occurs in this scenario with no effect for brief stimuli such as Von Frey filament stimulation but significant effect for prolonged stimuli such as active gait. Studies of midbrain receptor expression revealed certain key receptors including high early serotonin receptor (5HT2B) expression at 1 week, persistent high metabotropic glutamate receptor (mGluR5) at 1 and 4 weeks, and late and high expression of mu-opioid receptor (μ-OR) at 4 weeks.
The short-lived heightened 5HT2b expression in the ventral midbrain parallels findings for altered gait, where changes were observed at 1 week that returned to control levels by 4 weeks after surgery. Serotonin receptor modulation has been the subject of some study for mediating pain and nerve damage in lumbar radiculopathy [31], as well as for other incidences of nerve damage. Curiously, 5HT2b receptor polymorphisms have been associated in population studies with drug-seeking behavior, and also are found in humans to mediate the effects of amphetamines (MDMA). Following compression of the nerve root or cauda equina, serotonin has been found to constrict blood flow to the nerve root and to potentially contribute to sustained nerve injury [32, 33]. Administration of a specific serotonin receptor antagonist (5HT2) has been found to maintain blood flow levels and to eliminate the observed blood vessel diameter constriction. A pattern for increased serotonin receptor expression in the midbrain at short periods after injury may be consistent with a short-term nerve damage response, although additional studies would be required to determine if these central system changes could act to produce local changes in blood flow or other parameters at the site of nerve injury. Systemic delivery of serotonin 2 receptor antagonists have been shown to decrease allodynia [34] and have been used to treat pain and nociception in human subjects presenting with sciatica [32]. Based on prior studies showing that increased release of serotonin can occur in platelets and mast cells of injured or inflamed tissues that lead to hyperalgesic reactions in peripheral tissue [35], it is possible that an increased serotonin potentiation due to either elevated receptor expression or tissue injury contributes to the abnormal gait response observed in rats 1 week after surgery.
Post surgery, increased expression levels ofmetabotropic glutamate receptor 5 (mGluR5)occurred early at 1 week and persisted for4 weeks in the ventral midbrain. This pattern is consistent with the known roles of both the PAG and the dorsal midbrain raphe nuclei in pain perception and sensory function. A prior study reported an ability for mGluR5 antagonists to reduce allodynia and hyperalgesia following spinal nerve ligation [36-38] further confirming a role for this receptor in mediating sensitivity. In addition, animal models of neuropathic pain have demonstrated an increase in glutamatergic input from primary afferents to spinal dorsal horn neurons after injury, resulting in increased excitation of mGluR receptors [39, 40]. The observed increase in mGluR5 receptor expression levels provides evidence of potential supraspinal altered glutamatergic transmission in the setting of peripheral nerve injury. Importantly, the contradistinction with early increased mGluR5 levels in the sham animals, and subsequent reversal to control levels at 4 weeks after surgery, clearly demonstrates that placement of NP tissue on the DRG is related to a differential and sustained effect that differs from that due to surgery alone.
Activation of the mu-opioid receptor at the level of the midbrain PAG is known to decrease afferent noxious transmission by decreasing dorsal horn neuron firing, decreasing nociceptive transmission through a physiology involving decreased potassium conductance. Stimulation of this region in animal models of acute pain has been demonstrated to alleviate allodynia and provide for phenotypic normalization, however no long-term studies have been performed to observe alteration of system sensitivity by changed receptor expression levels. All changes in the expression of the μ-opioid receptor following sham surgery or NP-treatment were noted in the ventral midbrain, likely primarily in the PAG, consistent with their anatomic localization to this region. In prior studies of nerve ligation, μ-OR expression has decreased (rather than increased) in the spinal cord following injury [41], while studies of chronic constriction nerve injury present findings for an increase in μ-OR in the spinal dorsal horn [42]. These observations suggest that μ-OR reactions depend on the mechanism of injury and more specifically on the integrity of the primary sensory neuron. Our findings of elevated μ-OR expression in the midbrains of both sham surgery and NP-treated animals at 1 week after surgery provide further evidence that μ-OR remodeling may be a broader response to local injury, but that the sustained and elevated expression of μ-OR in NP-treated animals at the 4 week timepoint may represent an adaptive response to how the animals overcome sustained allodynia to provide for normalization of gait.
A new finding here is the trend for elevated monocyte chemoattractant protein (MCP-1) in cerebrospinal fluid of NP-treated rats at 4 weeks after surgery. MCP-1 recruits monocytes to sites of injury and infection, and is elevated in the CSF of patients with significant spinal cord injury [43, 44]. These studies provide evidence that cord injury can be associated with modified profiles in the CSF for MCP-1 in addition to a number of other chemokines reported previously including interleukins-8 and -6. Nevertheless, epidural lavage fluids obtained from patients with disc herniation demonstrated no measurable differences in MCP-1 levels, or numerous other chemokines and cytokines, between fluids from normal volunteer subjects and affected patients [45]. While there is great interest in identifying a biomarker of radiculopathy-associated pathology, these results for MCP-1 are insufficient to suggest its utility as a measured biomarker for use in human subjects.
CONCLUSION
In summary, the results of this study identify sustained mechanical allodynia, yet transient changes in stance symmetry and gait, for rats following NP placement upon the lumbar DRG as a model of radiculopathy. Activated microglia at the DRG was also observed in rats of the radiculopathy model. Evaluation of changes in neurotransmitter receptors at the midbrain PAG suggests an adaptive response to radiculopathy that may alter sensitivity to endogenous activation. High levels of metabotropic glutamate receptor 5 persist over the course of the study while only at late time points does μ-opioid receptor show increased expression in the PAG region of the midbrain. Additional studies would be helpful to reveal the kinetics of midbrain receptor expression for a broader and more comprehensive panel of receptors following induction of radiculopathy using this rat model. These results are important for demonstrating that local injury at the DRG is associated with central remodeling at the midbrain, and points towards potential targets for therapeutic interventions for the treatment of radiculopathy.
AUTHORS’ CONTRIBUTIONS
All authors read and approved the final manuscript. PYH, MFS and LAS designed or performed the midbrain pain receptor immunostaining and methods for analysis. KDA designed, performed and analyzed the study of gait in these animals. MFS, LAS and WJR designed the surgical model and MFS led all animal surgeries with support from MAG. LJ, BAM, MAG and PYH designed or performed dorsal root ganglion immunostaining and methods for analysis. JLH and VBK acquired and analyzed data for cerebral spinal fluid biomarkers. WJR designed the surgical model and provided supervision of MFS and MAG in the completion of animal surgeries. LAS contributed to study design and funding, and supervised the study at all levels, oversaw data analysis and finalized the manuscript.
ACKNOWLEDGEMENTS
The authors would like to thank Mr. Stephen Johnson for his assistance with the surgical procedures.
Funding Support: This work was supported with funds from the NIH (P01AR050245, R01AR047442, and K99AR057426).
CONFLICT OF INTEREST
The authors confirm that this article content has no conflicts of interest.

