All published articles of this journal are available on ScienceDirect.

The Potential of N-Rich Plasma-Polymerized Ethylene (PPE:N) Films for Regulating the Phenotype of the Nucleus Pulposus
Authors Info & Affiliations
Abstract
We recently developed a nitrogen-rich plasma-polymerized biomaterial, designated “PPE:N” (N-doped plasma-polymerized ethylene) that is capable of suppressing cellular hypertrophy while promoting type I collagen and aggrecan expression in mesenchymal stem cells from osteoarthritis patients. We then hypothesized that these surfaces would form an ideal substrate on which the nucleus pulposus (NP) phenotype would be maintained. Recent evidence using microarrays showed that in young rats, the relative mRNA levels of glypican-3 (GPC3) and pleiotrophin binding factor (PTN) were significantly higher in nucleus pulposus (NP) compared to annulus fibrosus (AF) and articular cartilage. Furthermore, vimentin (VIM) mRNA levels were higher in NP versus articular cartilage. In contrast, the levels of expression of cartilage oligomeric matrix protein (COMP) and matrix gla protein precursor (MGP) were lower in NP compared to articular cartilage. The objective of this study was to compare the expression profiles of these genes in NP cells from fetal bovine lumbar discs when cultured on either commercial polystyrene (PS) tissue culture dishes or on PPE:N with time. We found that the expression of these genes varies with the concentration of N ([N]). More specifically, the expression of several genes of NP was sensitive to [N], with a decrease of GPC3, VIM, PTN, and MGP in function of decreasing [N]. The expression of aggrecan, collagen type I, and collagen type II was also studied: no significant differences were observed in the cells on different surfaces with different culture time. The results support the concept that PPE:N may be a suitable scaffold for the culture of NP cells. Further studies are however necessary to better understand their effects on cellular phenotypes.
INTRODUCTION
Intervertebral discs (IVDs) function as hydrostatic shock absorbers that cushion the load between adjacent vertebrae while maintaining mechanical coupling between them [1]. They are composite structures of the peripheral collagen-rich annulus fibrosus (AF) surrounding the proteoglycan-rich central nucleus pulposus (NP), which is sandwiched above and below by cartilage endplates [2]. The structure of the IVDs change with age, ultimately resulting in tissue degeneration [3, 4]. In the adult, the IVDs are characterized by their abundant extracellular matrix and low cell density [5], coupled with an absence of blood vessels, of a lymphatic system, and of nerves in all but the most peripheral annular layers [3]. The discs provide flexibility to the spinal column, allowing bending and torsion. They resist compressive forces by their high content of the proteoglycan aggrecan [6].
IVD degeneration is strongly associated with back pain [7]. While present management of disc pathology has been focused on symptoms associated with degeneration, fewer studies have so far been devoted to disc regeneration [8-10]. Although surgical procedures such as disc excision and vertebral fusion [11, 12] lead to relief of pain in the short term, they may also alter the biomechanics of the spine, leading to further degeneration of surrounding tissues and of discs at adjacent levels. The failure rates for lumbar fusions are typically 20% to 40% after five years [13].
As an alternative, research on tissue engineering of the IVD has recently been reported [8-10, 14-16]. The type of repair needed would certainly vary with the extent of IVD degeneration [17, 18]. Repair of existing tissue may be possible during early degeneration. Our approach is to inject cells, seeded in a supporting biomolecular scaffold, into the NP where they can be induced to produce an ECM rich in aggrecan [8, 9]. Several recent investigations have also shown that it may be possible to use mesenchymal stem cells (MSCs) for IVD repair and regeneration [19-24]. It is however difficult to imagine a benign site from which normal autologous disc cells can be harvested for implantation and it is therefore likely that it will be necessary to obtain such cells via stem cell differentiation: this at present is not a trivial task, as there is no cell marker that can be used to identify an authentic disc cell.
We recently showed that the production of an extracellular matrix with a high proteoglycan-to-collagen ratio can be used in vivo to distinguish NP cells from chondrocytes, and can help to identify an NP-like phenotype in vivo from a chondrocyte, when MSCs are induced to differentiate for tissue engineering of the disc [25]. We also developed a nitrogen (N)-rich plasma polymer biomaterial, designated "PPE:N" (N-doped plasma-polymerized ethylene) that is capable of suppressing cellular hypertrophy while promoting type I collagen and aggrecan expression in mesenchymal stem cells from osteoarthritis patients. We then hypothesized that these surfaces would form an ideal substrate on which the nucleus pulposus (NP) phenotype would be maintained.
Recent evidence from other laboratories also indicates that glypican-3 (GPC3), vimentin (VIM), pleiotrophin heparin binding factor (PTN), matrix gla protein (MGP), cartilage oligomeric matrix protein (COMP), and keratin 19 (KRT 19) are differently expressed in rat NP cells than are AF and articular cartilage cells [26], suggesting that these genes can be used as markers to distinguish NP cells from hyaline cartilage. GPC3 is a member of the glypican family, which contains a core protein anchored to the cytoplasmic membrane via a glycosyl phosphatidylinositol linkage. These proteins may play a role in the control of growth and cell division [27]. VIM is a member of the intermediate filament family of proteins that plays a significant role in supporting and anchoring the position of the organelles in the cytosol [28]. PTN is an 18-kDa growth factor that has a high affinity for heparin. It can upregulate proteoglycan synthesis in cartilage [29]. MGP is a calcium-binding protein found in numerous body tissues, including bone and cartilage. Its expression is upregulated in endochondral ossification, pathological calcification of cartilage, and hypertrophic chondrocytes during fracture healing [30, 31]. COMP is a noncollagenous extracellular matrix protein containing five identical glycoprotein subunits, each with calcium-binding (thrombospondin-like) domains. Its binding to other ECM proteins, such as collagen, appears to depend on divalent cations [32]. KRT 19 is a member of the keratin family that are intermediate filament proteins responsible for the structural integrity of epithelial cells. KRT 19 was found to be expressed in the NP together with VIM [33].
The objective of this study was to compare the expression profiles of NP cells from fetal bovine lumbar discs when cultured on either commercial polystyrene (PS) tissue culture dishes or on PPE:N with time.
MATERIALS AND METHODS
Deposition of PPE:N
PPE:N films were deposited on biaxially-oriented polypropylene (BOPP, graciously provided by 3M Company), in the form of a 50 µm-thick isotactic polymer film [34, 35], as previously described [36]. Our atmospheric pressure plasma deposition system comprised a moveable aluminium plate electrode, on which flat sheets of BOPP (typically 22 cm x 30 cm) were uniformly coated in a 10 kHz, high voltage (~ 18 kV peak-to-peak) dielectric barrier discharge (DBD) [6]. This was accomplished by carefully-controlled linear displacement of the substrate through the ca. 1 cm wide discharge zone between the cylindrical, ceramic-coated high voltage electrode and the grounded aluminium plate, spaced 1 mm apart. Using this apparatus, films containing various nitrogen concentrations, [N], were deposited from the precursor gas mixture composed of very pure nitrogen (N2, 10 standard liters per minute, slm) and ethylene (C2H4, typically some tens of standard cubic centimeters per minute, sccm).
PPE:N Films with Different Nitrogen Concentrations ([N])
Obtaining PPE:N films with different [N] values was accomplished as previously described [36, 37]. Briefly, the flow rate (F) of the ethylene gas precursor, FC2H4, was changed from the lowest value of 5 sccm to higher values, thereby gradually reducing [N]. Thus, for FC2H4 = 5 sccm, [N] ≈ 36%, while for the highest FC2H4 used here, 60 sccm, [N] ≈ 17%: all other deposition conditions of course being maintained constant. Table 1 shows the characteristics of the three different PPE:N coatings we have used in the present investigation. Hereafter in this text, we shall refer to these PPE:N coatings in terms of the FC2H4 values used in their preparation, namely 5, 10, and 60 sccm, knowing that the resulting different [N] values can lead to significantly different cell responses [36, 37]; accordingly, as shown in Table 1, coatings are designated S5, S10, and S60, in the order of increasing FC2H4 (decreasing [N]).
Characteristics of PS Control and PPE:N Surfaces
| Sample | Nitrogen Flow Rate, FN2 (slm) | Ethylene Flow Rate, FC2H4 (sccm) | Elemental Concentrations (at %) | ||
|---|---|---|---|---|---|
| Nitrogen [N] | Oxygen [O] | Carbon [C] | |||
| S5 | 10 | 5 | 36.0 | 9.0 | 55.0 |
| S10 | 10 | 10 | 29.5 | 7.0 | 63.5 |
| S60 | 10 | 60 | 18.0 | 5.0 | 77.0 |
| PS | ---- | ---- | 0 | 18.0 | 82.0 |
The surface compositions of the thin PPE:N films were determined by X-Ray photoelectron spectroscopy (XPS) using a Kratos Axis Ultra system, with non-monochromatic Mg Kα radiation [34]; at least five samples were analysed for each condition. We will be referring to their surface-elemental concentrations, [X], in terms of the elements that comprise PPE:N, namely N, C and O; since hydrogen cannot be detected by XPS, [X] is given by:
N, O, and C being determined from XPS broadscan spectra. Other methods used for the characterization of PPE:N films included attenuated total internal reflectance Fourier transform infrared spectroscopy (ATR-FTIR), contact angle goniometry, atomic force microscopy, stylus profilometry and spectroscopic ellipsometry, as described elsewhere [34].
Source of Intervertebral Discs (IVDs)
Three fetal bovine lumbar discs were obtained from the slaughterhouse Les Abattoirs Colbex-Levinoff (St-Cyrille de Wendover, QC). Ages of the fetal bovine were determined by the method of Pal et al. based on the length of the tibia [38]. The ages of the fetal bovine are about 5 months old. IVDs were classified as nondegenerated grade I according to the grading system of Thomson [39].
Cell Isolation
Cells were isolated immediately after transportation from the slaughterhouse (abattoir). The IVDs were dissected from their adjacent vertebral bodies, placed in DMEM-high glucose containing 150 ng/ml gentamycin, 100 U/ml penicillin, 100 µg/ml streptomycin, and 0.25 µg/ml fungizone (medium A). Under aseptic conditions, the IVDs were separated by dissection into regions corresponding to the AF and the NP. The NP regions were, in turn, cut into approximately 2 mm fragments and were washed twice in medium A for 15 min. Cells were enzymatically isolated from the tissue, using the method adapted from protease type XIV/collagenase protocol [9]. Briefly, the tissue was washed and then digested overnight at 37°C in washing medium (medium A, without gentamicin and fungizone) supplemented with 10% fetal bovine serum (FBS, Hyclone) and bacterial collagenase type IA (Sigma-Aldrich, Oakville, ON) at 0.03% (w/v). The resulting cell suspensions were passed through a 70-µm cell strainer (Beckton Dickinson, Oakville, ON), washed twice in DMEM-high glucose containing 10% FBS, 100 U/ml penicillin, and 100 µg/ml streptomycin. Cells were counted with a hemacytometer and the viability was determined using 0.04% Trypan Blue Dye. Approximately 5 millions cells from 10 g of the NP per fetus were recovered and used.
Cell Culture
NP cells were cultured on each of the three different PPE:N coatings on BOPP in DMEM-high glucose supplemented with 10% FBS, 100 U/ml penicillin, and 100 µg/ml streptomycin. At the beginning about 1.0 million cells were layered on S5 surfaces tightly covering 100 x 20 mm culture dishes. Commercial polystyrene (PS) tissue culture dishes (Sarstedt, Montreal, QC) were used as controls (hereafter designated “PS control”). These commercial culture dishes are known to be plasma-treated for improved hydrophilicity and cell-adhesion, and our XPS measurements show that they possess a surface oxygen concentration, [O], of 18%. In all cases, the medium was changed every 2 days. The cells were then harvested at day 3 and day 14 to study the short- and long-term effect, respectively, of surfaces on gene expression.
Total RNA Isolation
Total RNA was extracted from NP cells by a modification of the method of Chomcynski and Sacchi [40] using TRIzol reagent (Invitrogen, Burlington ON). The aqueous phase was precipitated in 1 volume of isopropanol. The resulting RNA pellet was air-dried, resuspended in 50 µl of diethylpyrocarbonate-treated water, and 10 µl was assayed for RNA concentration and purity by measuring A260/A280.
Reverse Transcription (RT) and Polymerase Chain Reaction (PCR)
The RT reactions were performed using 1 µg total RNA isolated from the NP cells in a total volume of 20 µ1, containing 50 mM Tris-HCl (pH 8.3), 75 mM KCl, 3 mM MgCl2, 10 mM DTT, 50 µM each of dATP, dGTP, dCTP and dTTP, and 200 units of Superscript II - RNase H reverse transcriptase (Invitrogen).
PCR was performed in a total volume of 25 µl containing: 10 mM Tris-HCl (pH 8.3), 1.5 mM MgCl2, 0.4 mM of dATP, dGTP, dCTP, dTTP, 0.8 µM of each primer, 1 µl of RT mixture and 2.5 units of Taq DNA polymerase (Invitrogen) as previously described [40]. The 25 cycles of PCR (except GPC3 with 35 cycles) included denaturation (94°C, 1 min), annealing (56°C, 45 sec) and extension (72°C, 40sec). The PCR condition was applicable to all genes. After agarose (2%) gel electrophoresis, PCR products were visualized by ethidium bromide staining and analyzed using a Bio-Rad VersaDoc image analysis system, equipped with a cooled 12 bit CCD camera (Bio-Rad, Mississauga ON). GAPDH (Glyceraldehyde-3-phosphate dehydrogenase) was used as housekeeping gene. To confirm the absence of chromosomal DNA contamination of RNA samples, PCR was also performed with RNA aliquots. The primers were obtained from InVitrogen. Their sequences, shown in Table 2, were chosen because they are specific for bovine genes and amplify a unique product.
Primers Used for RT-PCR Analyses
| Gene | Primers | Size |
|---|---|---|
| GPC3 | 5’-ACAGCACGATTGAACATGGA -3’ 5’-TTGATGTCAGAACCCAGGATG -3’ |
236bp |
| VIM | 5’-TGCAGGATG GA TTCAGAACA -3’ 5’-ATCGTGATGCTGGGAAGTTT -3’ |
296bp |
| PTN | 5’-TGACTGTGGAGAATGGCAGTG -3’ 5’-TTGCCACAGGGCTTG GAG AT -3’ |
294bp |
| MGP | 5’-TCCTTCTCTCCATCCTGGCT -3’ 5’-AAATAACGGTCGTAGGCAGCA -3’ |
274bp |
| COMP | 5’-ATGACGACTACGCTGGTTTCA -3’ 5’-TGGTGTCGTTGCAGCGGTA -3’ |
456bp |
| AGG | 5’-CAGAACATGCGCTCCAATGA -3’ 5’-CGTCATAGGTTTCGTTGGTG -3’ |
370bp |
| COL1A2 | 5’- GGTCACAATGGTCTGGATGGA -3’ 5’- TTGTTGCCGCTCTCTCCTTT-3’ |
363bp |
| COL2A1 | 5’-GAACCCAGAAACAACACAATCC-3’ 5’-GAGGGGAGAAAAGTCCGAAC-3’ |
168bp |
| GAPDH | 5’-TATGACCACTGTCCACGCCAT -3’ 5’-AGTATCGCTGTTGAAGTCGCA -3’ |
349bp |
GPC3: glypican-3, VIM: vimentin, PTN: pleiotrophin, MGP: matrix Gla protein, COMP: cartilage oligomeric matrix protein, AGG: aggrecan, COL1A2: collagen type I alpha 2, COL2A1: collagen type II alpha 1, GAPDH - glyceraldehyde-3-phosphate dehydrogenase.
Statistical Analysis
Data are presented as the mean ± standard error of 3 experiments. Statistical differences were calculated using Student t-test, with p < 0.05 as the level of significance.
RESULTS
Characteristics of PPE:N Coatings
In an earlier article, we reported in considerable details the methodology for depositing PPE:N coatings on BOPP and on other (polymeric or glass) substrates, as well as methods used for characterizing the resulting thin film materials, and the biological responses, mostly adhesion or non-adhesion of various cell lineages of interest to these authors [36]. In order not to unduly repeat those earlier-published data, we have limited ourselves here to describing how [N] varies with FC2H4. As shown earlier [36], [N] decreases in a non-linear monotonic manner from the maximum value, [N] ≈ 36% for the lowest value of FC2H4 (5 sccm; S5), to [N] ≈ 17% for the highest FC2H4 value used (60 sccm; S60) (Table 1). In this article, we limit our characterization of different PPE:N substrates to their elemental composition, in particular their total [N]; however, more realistically, the substrates’ effect on adhering cells is mediated by the concentrations of various chemical functionalities at the surface, for example amines, imines, nitriles, amides, acids and alcohols (bound oxygen is always incorporated in plasma polymer films due to the reaction of residual surface radicals with ambient air). Unfortunately, plasma polymers are difficult to characterize, on account of their random highly cross-linked structure; the resulting peak broadening, therefore, greatly complicates quantitative, even qualitative, analysis by most spectroscopic methods (high resolution XPS, FTIR, solid state NMR, etc.), and this inherently limits our knowledge of the specific surface chemistries of PPE:N films at the present time. However, as reported earlier [34], we do know (from chemical derivatization experiments, coupled with XPS, and from FTIR measurements) that primary amines account for 5 to 10 % of [N], and that nitriles (-C≡N) also constitute an important surface functionality; these results are in keeping with surface-analytical experiments on these same materials now in progress, using Near-Edge X-Ray Absorption Fine-Structure (NEXAFS), to be reported elsewhere. Primary amines are positively charged, and their particular efficacy is believed to result from the fact that they attract negatively charged proteins and other bio-molecules from the culture medium; alternately, proteins secreted from the cells’ own ECM preferentially form covalent bonds with these favored reactive amine sites.
Cells from the Fetal Bovine Lumbar Spine
Fig. (1) shows the microscopic aspect of NP cells in culture. When cultured on PS control, NP cells were able to grow and reach confluence after 14 days (Fig. 1A); at low [N] (sample S60), cell growth was not affected (Fig. 1B) while cell proliferation was decreased on S5 surfaces (Fig. 1D). Quantitatively, cell number increased by 2.3 times on the PS control over the 14 days of culture while this increase was of 2.2, 2.0, and 1.6 times on S60, S10, and S5 surfaces, respectively, suggesting a dose-dependent effect of N on NP cell proliferation.

Culture of NP cells from fetal bovine lumbar discs on N-rich plasma polymer layers. NP cells from ~ 5 months old fetal bovine lumbar discs were cultured on control PS control (A), S60 (B), S10 (C), and S5 (D) surfaces.
Expression of Glypican-3 (GPC3)
The results of RT-PCR analyses of GCP3 gene expression, normalized to GAPDH expression, are shown in Fig. (2). The expression of GPC3 decreased with time on PS controls. Its expression was significantly reduced in the NP cells cultured on the S60 surface after short (3 days) and long (14 days) culture time. Interestingly, there was less downregulated of GPC3 expression when cells were cultured for 14 days on the S5 layer.
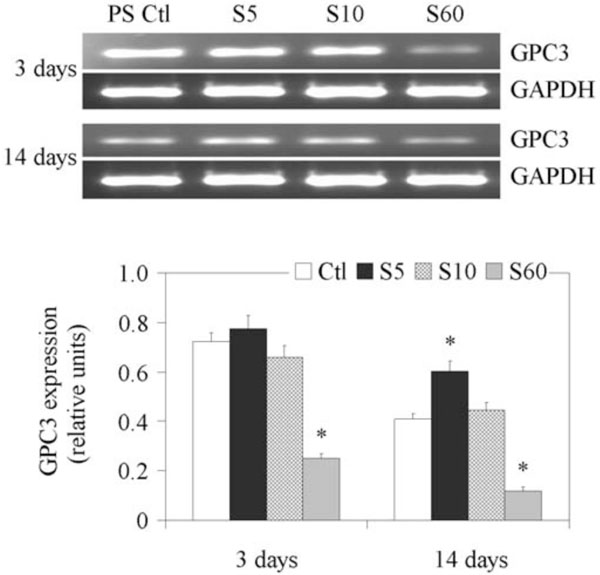
Expression of GPC3 gene in fetal bovine NP cells. NP cells from lumbar discs of ~ 5 months old fetal bovines were cultured for up to 14 days on PS control and different PPE:N surfaces. GAPDH was used as housekeeping gene and served to normalize the results. Agarose gels are representative of three experiments. Semi-quantitative results are the mean ± standard error of these three experiments. *p < 0.05 vs PS control.
Expression of Vimentin (VIM)
Since VIM has been shown to be expressed at significant levels in the NP [26], we next explored whether this message could be regulated by PPE:N surfaces in fetal bovine NP cells. Unlike the GPC3 message (Fig. 3), the expression of VIM was maintained throughout the culture when cells were cultured on PS controls (Fig. 3).
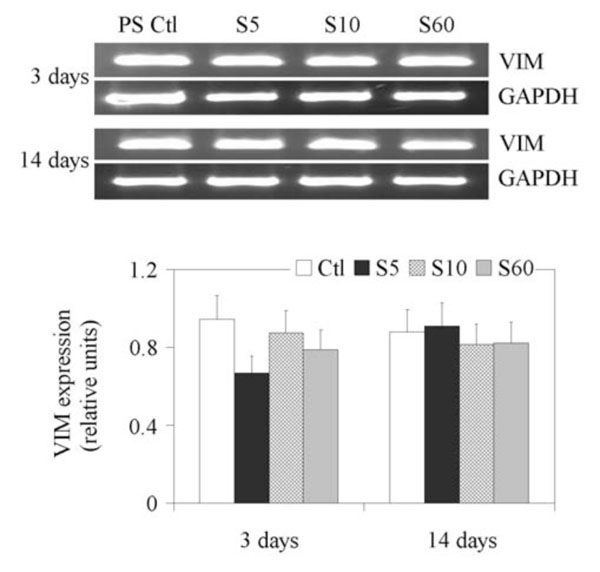
Expression of VIM gene in fetal bovine NP cells. NP cells from lumbar discs of ~ 5 months old fetal bovines were cultured for up to 14 days on PS control and different PPE:N surfaces. GAPDH was used as housekeeping gene and served to normalize the results. Agarose gels are representative of three experiments. Semi-quantitative results are the mean ± standard error of these three experiments.
Expression of Pleiotrophin Heparin Binding Factor (PTN)
Since PTN was recently discovered to be expressed in more than 10-fold higher concentration in the NP than in the AF [26], we next explored its expression on the different culture surfaces. Results are shown on Fig. (4). The expression of PTN on the PS controls did not change significantly from day 3 to day 14, but decreased when cultured on all three PPE:N coatings on day 3 and day 14. The weakest expression of PTN was observed on day 3 when NP cells were cultured on S60.
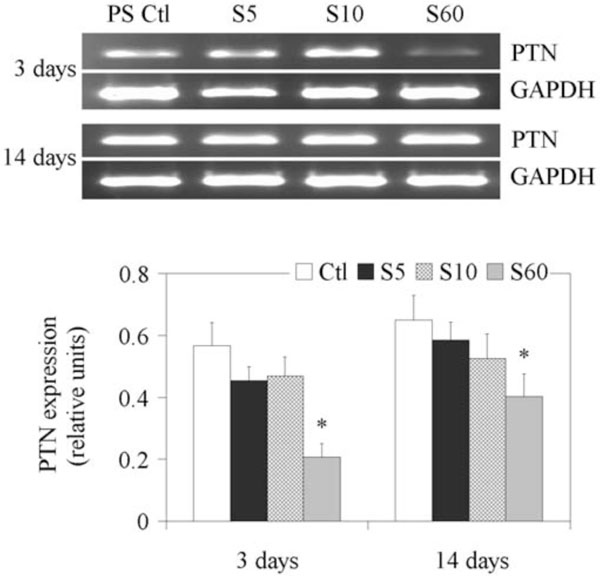
Expression of PTN gene in fetal bovine NP cells. NP cells from lumbar discs of ~ 5 months old fetal bovines were cultured for up to 14 days on PS control and different PPE:N surfaces. GAPDH was used as housekeeping gene and served to normalize the results. Agarose gels are representative of three experiments. Semi-quantitative results are the mean ± standard error of these three experiments. *p < 0.05 vs PS control.
Expression of Matrix Gla Protein (MGP)
The results of RT-PCR analyses of gene expression of MGP are shown in Fig. (5). The expression of MGP on PS control was higher on day 3 compared to day 14. Its expression was decreased when NP cells were cultured on all three PPE:N surfaces on day 3 and day 14. Similarly to PTN, its expression decreased with decreasing [N].
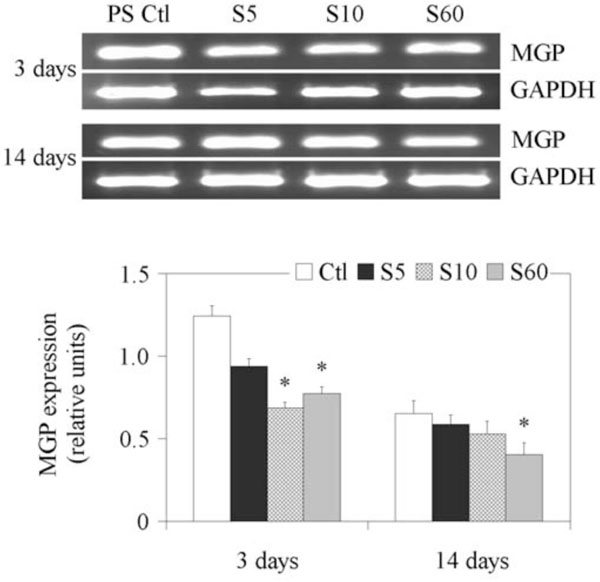
Expression of MGP gene in fetal bovine NP cells. NP cells from lumbar discs of ~ 5 months old fetal bovines were cultured for up to 14 days on PS control and different PPE:N surfaces. GAPDH was used as housekeeping gene and served to normalize the results. Agarose gels are representative of three experiments. Semi-quantitative results are the mean ± standard error of these three experiments. *p < 0.05 vs PS control.
Expression of Cartilage Oligomeric Matrix Protein (COMP)
The results of RT-PCR analyses of COMP gene expression are shown in Fig. (6). Its expression was consistently detectable in NP cells cultured on PS controls from day 3, following which its expression decreased progressively and significantly with time. A decrease was observed when cells were cultured on S5, S10 and S60 surfaces on day 3, but the different PPE:N surfaces had no significant effect on COMP expression on day 14.
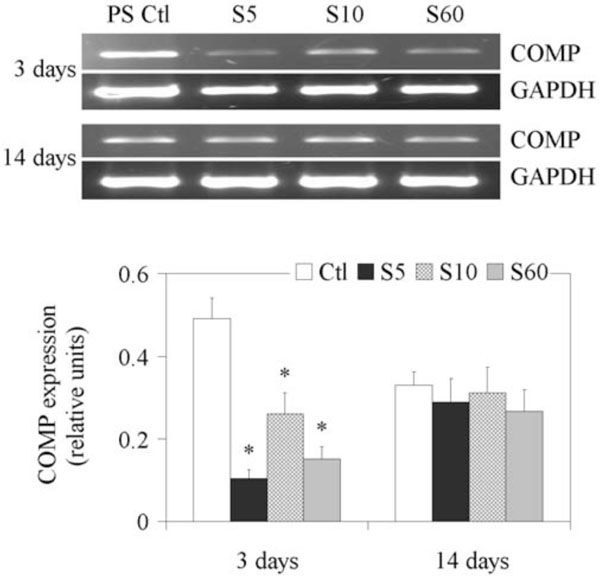
Expression of COMP gene in fetal bovine NP cells. NP cells from lumbar discs of ~ 5 months old fetal bovines were cultured for up to 14 days on PS control and different PPE:N surfaces. GAPDH was used as housekeeping gene and served to normalize the results. Agarose gels are representative of three experiments. Quantitative results are the mean ± standard error of these three experiments.*p < 0.05 vs PS control.
Expression of Aggrecan, Collagen Type I, and Collagen Type II
We finally looked at the effect of PPE:N surfaces on genes of the extracellular matrix of NP cells from fetal bovine lumbar discs. Aggrecan (AGG), collagen type I alpha 2 (COL1A2), and collagen type II alpha 1 (COL2A1) gene expression stayed stable throughout the culture time. There was also no effect of the PPE:N surfaces on their expression (results not shown).
DISCUSSION
Tissue engineering involves the interaction of cells with a material surface, by way of protein adsorption from the medium, or that secreted by the cells’ own extracellular matrix [8, 9]; therefore, the nature (composition, morphology) of the surface can directly influence various cellular responses [34, 41-46]. The attachments made by the cells to their substrate can also determine specific gene expression profiles related to cellular phenotypes [43, 44]. While some studies report on the phenotype of intervertebral disc (IVD) cells [24, 25, 47, 48], little is known of how their phenotype can be affected by substrate surfaces. One of the major obstacles to understanding the role of material surfaces in regulating the responses of disc cells has been the lack of biocompatible substrates with chemically-bound functionalities that can readily be controlled. The recent advances in our group in creating novel bioactive synthetic polymer surfaces, for example, plasma deposition of the present PPE:N films, enabled us to study the culture of disc cells, and of other cell types, on these new surfaces [34, 37]. The regulation of the NP phenotype is clearly a complex process. Its examination in culture offers an opportunity to gain a better understanding of the different molecular changes and regulatory mechanisms that are involved in NP maturation and degradation. The present RT-PCR analyses showed that genes expressed in NP cells from fetal bovine lumbar disc are expressed differently when cells were cultured on different PPE:N surfaces. Indeed, both the culture time and the [N] were found to change the expression of most of the genes we studied in these cells. To the best of our knowledge, no work presented to date has addressed the question whether synthetic materials similar to our N-rich plasma-polymers could possibly offer a substrate suitable for the regulation of the disc phenotype. A further impetus for the present work has been the finding that the markers previously reported in rat NP [26] and dog notochordal [37] cells are also expressed in their bovine NP cells.
Importantly, the present results show that it is possible to maintain the expression of the reported potential markers of NP phenotype throughout the period of culturing NP cells on PS controls. The expression of AGG, COL1A2, and COL2A1, which are important components of the extracellular matrix, was also maintained throughout the time of culture. This was surprising as we expected type II collagen to be downregulated and type I collagen upregulated when the cells were cultured on plastic. These results indicate that the cells’ phenotype can be maintained or is only mildly affected for at least 14 days in culture and there was no cell division. However, polystyrene cannot be used as a scaffold for tissue engineering of the disc, and alternatives have to be investigated. Our results suggest that N-rich surfaces are good candidates to maintain the expression profile of NP cells from fetal bovine lumbar discs.
GPC3 is a member of the glypican family of heparin sulfate proteoglycan. In general, glypicans are expressed predominantly during development and its expression levels change with age [49]. In the present study, we demonstrated that GPC3 is expressed in fetal bovine NP cells. The expression of VIM, a cytoskeletal protein, was maintained throughout the time in culture and was not affected significantly by the different PPE:N surfaces. A difference in the expression pattern on PPE:N surfaces was observed for PTN, a secreted heparin binding cytokine, and for MGP, an extracellular matrix protein of the mineral-binding Gla protein family. The expression of COMP, a member of the thrombospondin gene family found in the extracellular matrix, was decreased in the cells cultured on PPE:N surfaces, suggesting a gene-specific effect of these surfaces. Similarly, the selective inhibition of gene expression when cells were cultured on PPE:N surfaces was observed in our earlier studies of the differentiation of mesenchymal stem cells [37, 43, 44, 50] and notochordal cells from spines of dogs [37]. Thus, inhibitory mechanisms that are dependent on [N] at the surface of the culture medium exist, whereby certain genes are selectively suppressed. This mechanism of interaction linking PPE:N surfaces and NP cells is however unknown. Preliminary results using MSCs and macrophages suggest that this may be cell specific (personal unpublished data). This remains to be investigated.
CONCLUSION
The present study shows that genes expressed in NP cells from fetal bovine lumbar discs are differently expressed when cells are cultured on PPE:N surfaces, suggesting a gene-specific effect of these surfaces on cellular behavior. Our results also suggest that [N] can affect NP cell proliferation. However, further studies are necessary to better understand their effects on cellular phenotypes.
ACKNOWLEDGEMENTS
This work is funded by grants from the Canadian Institutes for Health Research (CIHR), AO Foundation, Switzerland (to F.M and J.A), and from the Natural Sciences and Engineering Research Council of Canada (NSERC) (to M.R.W).

